En el anterior artículo hablamos de cómo algunos famosos lagartos se pusieron a dos patas para cumplir sus sueños. Bien, con este otro artículo desvelamos que su verdadera intención no es otra que la de seguir viviendo. Si unimos la capacidad de correr a dos patas con la que nombraremos a continuación, dejamos claro que, si no eres un poderoso cocodrilo o una resistente tortuga, eres un cobarde escamoso, que poco más puede hacer que engañar con el rabo.
Muchos lagartos pueden soltar una parte de su cola en respuesta al ataque de un depredador, un comportamiento conocido como autotomía caudal. Debido a que no se conoce ese comportamiento en cocodrilos y tortugas, se sugiere que la capacidad evolucionó en la rama Lepidosauria (tuátaras, lagartos, serpientes y anfisbenios).
La autotomía caudal
La autotomía caudal de Squamata, o escamosos (lagartos, camaleones, iguanas y afines, serpientes y culebrillas ciegas), y la posterior regeneración de la cola se han estudiado extensamente debido a la importancia adaptativa de este comportamiento único antipredatorio (Etheridge, 1967; Arnold,1984; Seligmann et al., 2008; Bateman & Fleming, 2009; Clause & Capaldi, 2006). Además de aumentar las posibilidades de supervivencia al facilitar el escape, puede distraer a los depredadores y minimizar la pérdida de sangre después de una lesión (Bely & Nyberg, 2010). Los lagartos existentes muestran dos tipos de autotomía caudal:
- Autotomía intravertebral:Las vértebras caudales pueden separarse a lo largo de un plano de fractura preexistente que pasa a través del cuerpo vertebral y del arco neural. Sólo se observa en los lepidosaurios existentes y se asocia con una regeneración más extensa de la cola (Etheridge, 1967).
- Autotomía intervertebral:La división ocurre entre vértebras caudales adyacentes (Arnold, 1984; Bateman & Fleming, 2009). Se encuentra en varias especies de Agama y en algunas serpientes, y parece haber evolucionado múltiples veces desde una condición ancestral no autotómica (Etheridge, 1967; Arnold,1984).
Ya que en este postnos vamos a centrar en la autotomía intravertebral, ahondemos en ello. La autotomía intravertebral ya se encuentra en las familias de lagartos filogenéticamente anteriores y el tuátara, con planos de fractura en sus vértebras caudales (Etheridge, 1967;Seligmann et al., 2008). Estos planos de fractura son áreas de debilidad a lo largo de la vértebra que resultan de la fusión incompleta de los esclerotomas (vértebras y discos intervertebrales, costillas y cartílagos costales) adyacentes durante el desarrollo vertebral (Winchester & Bellairs, 1977). En la vértebra totalmente osificada, el plano de fractura es un tabique transversal a través de la vértebra donde no se produce la deposición ósea.
Los captorínidos
Los únicos amniotas que muestran una evidencia convincente de autotomía caudal fuera de Lepidosauria son miembros del clado Captorhinidae (Price, 1940; Dilkes & Reisz, 1986). Los captorínidos, inicialmente, eran pequeños reptiles carnívoros (menos de 10 kg). Se originaron en el Carbonífero Tardío y se expandieron por todo el globo antes de la extinción en masa de finales del Pérmico (Müller & Reisz, 2005; Reisz et al., 2011; Ricqlès & Taquet, 1982), llenando numerosos nichos ecológicos como carnívoros, omnívoros y herbívoros pequeños, dentro de comunidades terrestres donde predominaban los depredadores (Reisz et al., 2015; LeBlanc et al., 2015; Reisz & Fröbisch, 2014; Modesto et al., 2014).
![]()
La importancia evolutiva de la autotomía caudal en reptiles paleozoicos nunca se ha discutido más allá de los informes preliminares sobre planos de fractura aparentes. Sin embargo, los avances en nuestra comprensión de la filogenia reptiliana ahora demuestran claramente que si los captorínidos realmente exhibían autotomía caudal, ésta fue la primera aparición de este comportamiento en Amniota, que más tarde evolucionó de forma independiente en los lepidosaurios, ya que los únicos registros de planos de fractura en las vértebras caudales de los captorínidos sugieren que no pasan completamente a través del cuerpo vertebral, como lo hacen en los lepidosaurios modernos (Etheridge, 1967; Arnold,1984; Price, 1940).
Además, es importante la posibilidad de una variación interespecífica u ontogenética a través de Captorhinidae, variando drásticamente el número de supuestas vértebras autotómicas en diferentes especies. Con ello se puede comprender lo extendido que estaba el comportamiento y si las presiones de depredación variaban, ocasionando dichos cambios en la capacidad de autotomía
Para abordar estos puntos, se ha examinado una muestra grande de vértebras caudales captorínidas aisladas, del sistema de cuevas kársticas de la localidad Richards Spur del Pérmico Temprano (Artinskiense, 289-286 millones de años), ubicada en Dolese Brothers Quarry en Oklahoma, EE. UU. Así, se compararon los supuestos planos de fractura en captorínidos con los de una iguana juvenil y otra adulta, ya que este escamoso moderno exhibe una reducción ontogenética de la autotomía caudal (Arnold,1984).
Las vértebras caudales de Captorhinidae y sus fracturas
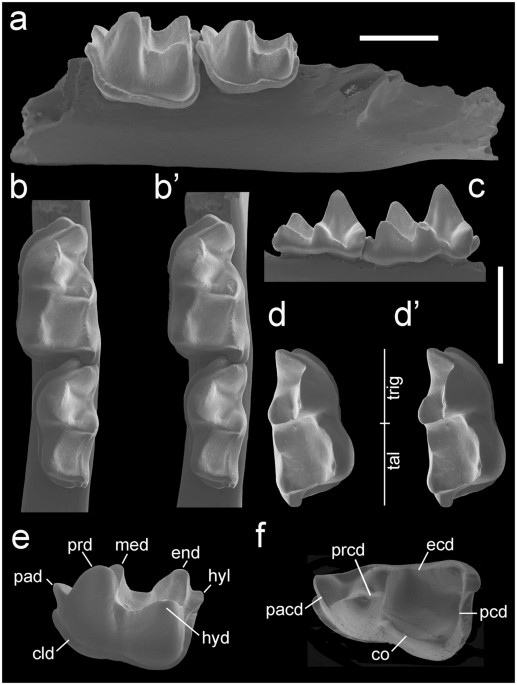
Estas vértebras caudales tienen divisiones transversales a lo largo de las superficies medio-ventrales del centro (Fig. 1a-c). Dichas divisiones no pueden ser características tafonómicas, ya que se encuentran consistentemente en la misma región de numerosos centros caudales y están ausentes en las vértebras caudales más anteriores, que contienen costillas (Fig. 1d). Las secciones delgadas longitudinales de vértebras caudales autótomas en captorínidos revelan que el plano de fractura se extiende a través de la pared ventral del cuerpo vertebral en forma de carrete y alcanza el suelo del canal notocordal a lo largo de la parte dorsoventral más estrecha del cuerpo vertebral (Fig. 1f, g).
![]() |
Fig. 1: Planos de fractura en vértebras caudales de captorínido.
(a) Reconstrucción artística del reptil pérmico Captorhinus con una cola autotómica (recuadro que muestra las vértebras caudales anteriores con planos de fractura). (b) Imagen y (c) imagen SEM de una vértebra caudal anterior aislada (ROM 73769) con un plano de fractura que pasa a través del cuerpo vertebral (flecha negra). (d) Vista ventral de una vértebra caudal anterior con soporte de costillas (ROM 77410) que muestra la ausencia de cualquier plano de fractura. (e) Vista ventral de una vértebra caudal con un plano de fractura (flechas negras) (ROM 73771) (f) sección delgada a través del plano sagital de una vértebra caudal (ROM 73773) con un plano de fractura (flecha negra) que pasa por la porción ventral del cuerpo vertebral. (g) Primer plano del plano de fractura (flechas negras) en (f) pasando al canal notocordal. Abreviaturas: cb, hueso cortical; cct, cartílago calcificado; ce, cuerpo vertebral; nc, canal neuronal; ns, espina neural; ntc, canal notocordal. Reconstrucción por Danielle Dufault. |
Ciertas comparaciones de las capas de tejido duro de cada cuerpo vertebral caudal captorínido, indican que la división se forma durante la osificación del cuerpo vertebral y no como resultado de la resorción ósea (eliminación de tejido óseo por los osteoclastos) a lo largo de la línea media del cuerpo vertebral. Los tabiques de autotomía se forman de forma similar durante la osificación de las vértebras caudales, no como en los escamosos modernos, por la resorción ósea (Winchester & Bellairs, 1977).
Las secciones transversales seriales a través de vértebras caudales que portan un plano de fractura confirman que estas características están restringidas a la mitad ventral de cada cuerpo vertebral. La brecha comienza en la base ventral de la vértebra, donde es considerable (Fig. 2a, b); atraviesa todo el ancho de las paredes laterales del cuerpo vertebral y hacia el canal notocordal (Fig. 2c, d), y se interrumpe en el hueso endocondral/endosteal que forma el núcleo interno de la vértebra (Fig. 2e, f). A pesar de estar restringido a la mitad ventral de cada cuerpo vertebral, los planos de fractura probablemente permitieran la autotomía caudal.
![]() |
| Fig. 2: Secciones transversales seriales a través del centro del plano de fractura de la vértebra caudal en captorínidos. (a) Sección transversal tomada cerca de la base del cuerpo vertebral (ROM 73771); (b) Primer plano del plano de fractura que atraviesa la superficie ventral del cuerpo vertebral en (a). (c) Sección transversal tomada al nivel del canal notocordal (ROM 73774). (d) Detalle del plano de fractura en (c) que pasa a través del cuerpo vertebral hacia el canal notocordal. (e) Sección transversal tomada a lo largo del techo del cuerpo vertebral (debajo del arco neural) (ROM 73774). (f) Detalle del plano de fractura en (g) que pasa a través del hueso cortical externo, pero no el hueso endoóseo/endocondral más interno. Abreviaturas: cb, hueso cortical; eb, hueso endocondral; rl, línea de inversión; sf, fibras de Sharpey. |
Una vértebra muestra una rotura aparente post-mortem a lo largo del plano de fractura, lo que proporciona una visión de la dinámica de la autotomía intravertebral en captorínidos. La vértebra muestra claramente un plano de fractura, basado en los bordes redondeados a cada lado del corte a lo largo de la superficie ventral del cuerpo vertebreal (Fig. 3). La ruptura posterior debe haber ocurrido algún tiempo después de la muerte y la desarticulación de los restos de este individuo. Sin embargo, la grieta sigue el plano de la fractura dorsalmente y se extiende ligeramente hacia la parte posterior hacia el extremo posterior del arco neural, rompiéndose las vértebras caudales con arcos neurales fusionados (Fig. 3).
![]() |
| Fig. 3: Trayectoria hipotética de rotura de autotomía en el centro caudal en los captorínidos. (a-c) Vistas lateral, anterior y ventral de un cuerpo vertebral caudal con plano de fractura, que muestra la extensión del plano de fractura, que no pasa por el arco o la espina dorsal (ROM 73774). (d, e) Imágenes SEM de una rotura post mortem en una vértebra caudal de un captorínido (ROM 77409) a lo largo de su plano de fractura. La ruptura sigue el plano de debilidad formado por el plano de autotomía (flecha blanca) y se encuentra en los lados izquierdo (d) y derecho (e), lo que sugiere que este fue el plano de debilidad en vida. (f) Reconstrucción de la autotomía caudal en una vértebra caudal. Durante la autotomía, la fractura sigue la ruta de menor resistencia a través de la mitad dorsal del cuerpo vertebral caudal y la base posterior del arco neural, evitando la espina neural. Abreviatura: na, arco neural; nc, canal neuronal; ns, espina neural; ntc, canal notocordal. |
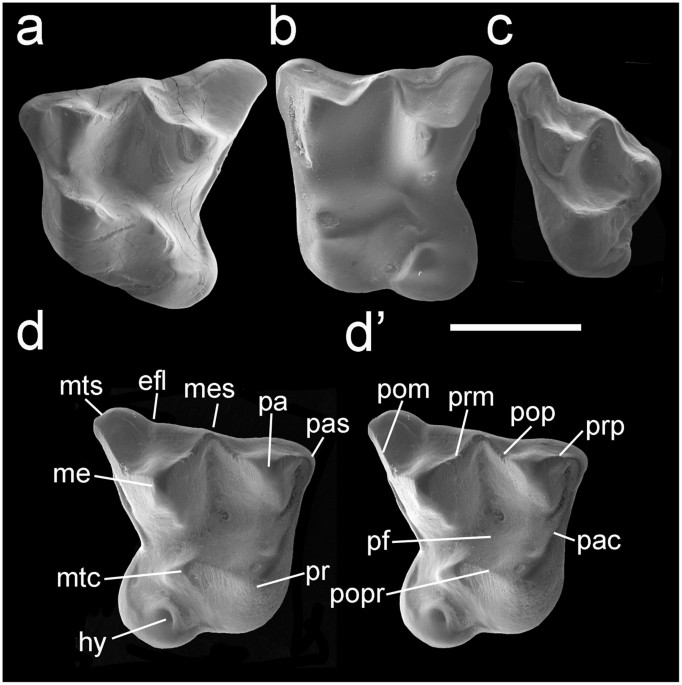
Se desconoce el número exacto de vértebras caudales en Captorhinus. Sin embargo, algunas vértebras se pueden asignar con confianza a las regiones anterior, media y posterior de la cola (Fig. 4a-c).
- Las espinas neurales de los caudales anteriores son altas y estrechas y se extienden postero-dorsalmente en ángulos relativamente altos. Sus respectivos centros son proporcionalmente cortos y biselados como resultado de la articulación con los arcos heméricos.
- Los caudales posteriores tienen un centro más largo y las espinas neurales se extienden a ángulos mucho más bajos por la parte postero-lateral (Fig. 4e-g). Estas vértebras medias-caudales frecuentemente poseen planos de fractura, lo que indica que una gran parte de la cola era autotómica.
Desafortunadamente, no se puede determinar el número exacto de vértebras autotómicas, pero sí se revelan al menos ocho vértebras de ese tipo, comenzando alrededor de la sexta a la octava caudal (Figs. 1a y 4a-c). Ocho vértebras caudales autóctonas es, por lo tanto, una estimación muy conservadora de los captorínidos de Richards Spur. Curiosamente, una serie caudal de un gran ejemplar de Captorhinus laticeps de la cantera McAnn de Oklahoma (Fig. 4c) muestra una sección autotómica mucho más corta de la cola, que está limitada a solo cuatro caudales anteriores.
![]() |
| Fig. 4: Variación en regiones caudales autótomas en captorínidos del Pérmico Temprano. (a) Serie caudal parcial de un captorínido juvenil (OMNH 03304) con al menos cinco vértebras autótomas, comenzando con la última caudal "process-bearing" (flecha). Tenga en cuenta la presencia de arcos neuronales sin fusionar. (b) Región autónoma de una serie caudal articulada (OMNH 1020, la imagen se ha recortado para incluir solo la serie caudal) con al menos ocho vértebras autóctonas, comenzando con la última caudal "process-bearing" (flecha). (c) Serie caudal parcial de Captorhinus laticeps (OUSM 15024, la imagen ha sido recortada para incluir solo la serie caudal) con una región autónoma que comienza en la octava caudal (flecha) y termina en la undécima (asterisco). (d) Reconstrucción de la región caudal conocida de Captorhinus (modificada desde) 22, que muestra la extensión de la región autónoma (coloreada) basada en (a-f). (e-g) Aislado anterior (proceso transversal), medio (sin procesos transversales, espinas neurales inclinados pronunciadamente) y posterior (espinas neurales de bajo ángulo) vértebras caudales de captorínidos de la localidad de Richards Spur. Abreviatura: tp, proceso transversal. |
Comparación con las vértebras caudales de iguana común
La anatomía y el desarrollo de las vértebras caudales autótomas en los escamosos se han descrito ampliamente (Etheridge, 1967; Winchester & Bellairs, 1977; Gilbert et al., 2013). Ahora compararemos las vértebras caudales con plano de fractura en captorínidos con vértebras autotómicas de especímenes juveniles y adultos de Iguana iguana (iguana común).
A diferencia de en los captorínidos, los planos de fractura en la iguana juvenil pasan a través de todo el centro y casi todo el arco neural (Fig. 5a-d).El plano de fractura está bordeado por hueso cortical compacto, similar a los planos de fractura en captorínidos.Las secciones longitudinales a lo largo de la línea media de las vértebras caudales muestran una capa delgada de hueso cortical en la extremidad dorsal del plano de fractura que parece unir las dos mitades del centro caudal (Fig. 1d). Durante la autotomía, esta división permite que la vértebra se separe, dejando una porción anterior corta del cuerpo vertebral y el arco neural (Fig. 5e). En el individuo más grande, el tabique de autotomía está cerrado por una masa de hueso cortical compacto y es apenas visible a lo largo de la superficie externa del centro (Fig. 5f-i).
![]() |
| Fig. 5: Comparaciones de planos de fractura a través de ontogenia en escamados y captorínidos. Las vértebras caudales en la iguana juvenil (ROM R9175) (a-d) tienen planos de fractura que se extienden a través de todo el cuerpo vertebral en sección delgada. Durante la autotomía, estos planos de fractura facilitan una ruptura en el tercio anterior de las vértebras, a través del arco neural y la columna vertebral, para permitir que la vértebra se divida en dos (e). Las iguanas más grandes y mayores (ROM 77408) (f-i) tienen planos de fractura cerrados y muestran una menor frecuencia de autotomía. Las grandes vértebras caudales anteriores de los captorínidos (ROM 73770) (j-m) poseen de forma similar planos de fractura cerrados en la sección delgada. Abreviaturas: cb, hueso cortical; nc, canal neuronal. |
Algunas vértebras caudales captorínidas muestran evidencia externa de un plano de fractura a lo largo de la superficie del cuerpo vertebral, pero el plano no pasa a través de la superficie ventral del hueso en una sección delgada (Fig. 5j-m). La región ventral media del centrum no es lisa, sino que posee una masa de hueso cortical mal organizado y vascularizado, lo que sugiere que, similar a la vértebra caudal más grande de la iguana, puede representar un plano de fractura cerrado.
Funcionamiento del mecanismo de autotomía y de regeneración
Los cortes tafonómicos en una vértebra caudal captorínida (Fig. 3) muestran que los planos de fractura pueden haber servido para propagar una fractura a lo largo de la base del arco neural cuando un depredador agarraba la cola. La fractura progresaría posteriormente a cualquier proceso transversal y al grueso de la espina neural (Fig. 3f). Esto hubiera permitido que un cuerpo vertebral caudal se dividiera en dos mitades desiguales durante la autotomía, incluso cuando los arcos neuronales se fusionaban con el centro. Esta forma de autotomía no habría dado lugar a una ruptura precisa a través de una sola vértebra, pero sin embargo sería efectiva para dejar caer la cola durante un ataque de un depredador.
Hasta la fecha, no hay evidencia que sugiera que los captorínidos pudieron regenerar una parte perdida de su cola.
Las diferencias en tamaño y grado de osificación de la muestra de vértebras autótomas sugieren que la capacidad de autotomizar la base de la cola es común a múltiples especies de captorínidos en la localidad de Richards Spur, y que había variabilidad ontogenética en la retención de la autotomía caudal. Desafortunadamente, la falta de esqueletos caudales completos para estas especies hace que sea difícil identificar qué especies captorínidas exhibieron autotomía caudal. Además, la naturaleza de los rellenos de cavernas y las operaciones de cantera en curso dificultan determinar el contexto estratigráfico de cualquier material recolectado del sitio (MacDougall et al., 2017; Kissel et al., 2002).
Además, este modelo, con el plano de fractura corriendo a través del centro, y luego pasando hacia atrás, hacia la espina neural, parece ser la explicación más parsimoniosa de cómo las vértebras pudieron separarse. Sin embargo, a diferencia de los lagartos, eso significaría que a pesar de dividir el centro por la mitad, todavía habría una conexión entre las zigapófisis de los arcos neurales. Incluso en captorínidos juveniles donde el arco neural no está fusionado con el centro, nuestro modelo requeriría una ruptura adicional entre las zigapófisis de las vértebras adyacentes. Por lo tanto, los captorínidos parecen tener un tipo único de autotomía caudal, una mezcla entre eventos intra e intervertebrales. La mayor parte de la ruptura ocurre a través del cuerpo del cuerpo vertebral caudal, parecido a la autotomía intravertebral, con la ruptura final ocurriendo en algún lugar entre las zigapófisis, posiblemente similar a la autotomía intervertebral.
Para concluir este post, resumimos que existe una amplia variación en las posiciones de los planos de fractura a lo largo o entre cada vértebra, así como en el número y las posiciones de las vértebras autóctonas (Etheridge, 1967; Pratt, 1946). En estos aspectos, los captorínidos fueron más similares a algunas iguanas y a los tuátaras existentes (Seligmann et al., 2008; Clause & Capaldi, 2006).
Muchos captorínidos pudieron soltar porciones significativas de sus colas. La frecuencia de pérdida de cola en los escamosos modernos se correlaciona fuertemente con la distancia de la región autónoma de la cloaca (Zani, 1996), lo que sugiere que el comienzo de la región vertebral autotómica cerca de la base de la cola puede reflejar una alta frecuencia de este comportamiento en poblaciones de captorínidos del Pérmico Temprano.
La autotomía caudal puede haber desempeñado un papel importante en la diversificación temprana de captorínidos, ya que fueron el primer grupo de amniotas en alcanzar una distribución casi global y diversificarse en nuevos nichos ecológicos en el Paleozoico Tardío (Reisz et al., 2011; Ricqlès & Taquet, 1982; LeBlanc et al., 2015; Modesto et al., 2014; Cisneros et al., 2015).
REFERENCIAS
- Etheridge, R. Lizard Caudal Vertebrae. Copeia 1967, 699 (1967).
- Arnold, E. N. Evolutionary aspects of tail shedding in lizards and their relatives. J. Nat. Hist. 18, 127–169 (1984).
- Seligmann, H., Moravec, J. & Werner, Y. L. Morphological, functional and evolutionary aspects of tail autotomy and regeneration in the ‘living fossil’ Sphenodon (Reptilia: Rhynchocephalia). Biol. J. Linn. Soc. 93, 721–743 (2008).
- Bateman, P. W. & Fleming, P. A. To cut a long tail short: a review of lizard caudal autotomy studies carried out over the last 20 years. J. Zool. 277, 1–14 (2009).
- Clause, A. R. & Capaldi, E. A. Caudal autotomy and regeneration in lizards. J. Exp. Zoolog. A Comp. Exp. Biol. 305A, 965–973 (2006).
- Bely, A. E. & Nyberg, K. G. Evolution of animal regeneration: re-emergence of a field. Trends Ecol. Evol. 25, 161–170 (2010).
- Winchester, L. & Bellairs, A. d’A. Aspects of vertebral development in lizards and snakes. J. Zool. 181, 495–525 (1977).
- Price, L. I. Autotomy of the Tail in Permian Reptiles. Copeia 1940, 119 (1940).
- Dilkes, D. W. & Reisz, R. R. The axial skeleton of the Early Permian reptile Eocaptorhinus laticeps (Williston). Can. J. Earth Sci. 23, 1288–1296 (1986).
- Müller, J. & Reisz, R. R. An early captorhinid reptile (Amniota, Eureptilia) from the Upper Carboniferous of Hamilton, Kansas. J. Vertebr. Paleontol. 25, 561–568 (2005).
- Reisz, R. R., Liu, J., Li, J.-L. & Müller, J. A new captorhinid reptile, Gansurhinus qingtoushanensis, gen. et sp. nov., from the Permian of China. Naturwissenschaften 98, 435–441 (2011).
- Ricqlès, A. de. & Taquet, P. La faune de vertebres de Permien Supérieur du Niger. I. Le captorhinomorphe Moradisaurus grandis(Reptilia, Cotylosauria)–le crane. Ann. Paléontol. 68, 33–63 (1982).
- Reisz, R. R., LeBlanc, A. R. H., Sidor, C. A., Scott, D. & May, W. A new captorhinid reptile from the Lower Permian of Oklahoma showing remarkable dental and mandibular convergence with microsaurian tetrapods. Sci. Nat. 102 (2015).
- LeBlanc, A. R., Brar, A. K., May, W. J. & Reisz, R. R. Multiple tooth-rowed captorhinids from the Early Permian fissure fills of the Bally Mountain Locality of Oklahoma. Vertebr. Anat. Morphol. Palaeontol. 1 (2015).
- Reisz, R. R. & Fröbisch, J. The Oldest Caseid Synapsid from the Late Pennsylvanian of Kansas, and the Evolution of Herbivory in Terrestrial Vertebrates. PLoS ONE 9, e94518 (2014).
- Modesto, S. P., Lamb, A. J. & Reisz, R. R. The captorhinid reptile Captorhinikos valensis from the lower Permian Vale Formation of Texas, and the evolution of herbivory in eureptiles. J. Vertebr. Paleontol. 34, 291–302 (2014).
- Gilbert, E. A. B., Payne, S. L. & Vickaryous, M. K. The Anatomy and Histology of Caudal Autotomy and Regeneration in Lizards. Physiol. Biochem. Zool. 86, 631–644 (2013).
- MacDougall, M. J., Tabor, N. J., Woodhead, J., Daoust, A. R. & Reisz, R. R. The unique preservational environment of the Early Permian (Cisuralian) fossiliferous cave deposits of the Richards Spur locality, Oklahoma. Palaeogeogr. Palaeoclimatol. Palaeoecol. 475, 1–11 (2017).
- Kissel, R. A., Dilkes, D. W. & Reisz, R. R. Captorhinus magnus, a new captorhinid (Amniota: Eureptilia) from the Lower Permian of Oklahoma, with new evidence on the homology of the astragalus. Can. J. Earth Sci. 39, 1363–1372 (2002).
- Pratt, C. W. M. The plane of fracture of the caudal vertebrae of certain lacertilians. J. Anat. 80, 184–188 (1946).
- Zani, P. A. Patterns of caudal-autotomy evolution in lizards. J. Zool. 240, 201–220 (1996).
- Cisneros, J. C. et al. New Permian fauna from tropical Gondwana. Nat. Commun. 6, 8676 (2015).



 † Indica taxón extincto; E, Early; L, Late; (#) , número de especies.
† Indica taxón extincto; E, Early; L, Late; (#) , número de especies.